

明日叶(Angelica keiskei)原产于日本,是一种食药兼用的蔬菜,属伞形科,为多年生林下草本植物,一直以来主要栽培于韩国、日本等地区[1].明日叶生长的适宜温度为15~22 ℃,喜阴,株高40~80 cm,外观与芹菜相似,茎秆呈圆柱形,内有黄色黏稠状液体;叶互生,三出羽状复叶,叶片呈深裂或浅裂掌状,细锯齿状边缘;5-10月开花,顶生复伞形花序,花呈乳白色.根据植株外观,可分为红茎种、青茎种和混合种[2].明日叶是一种嫩茎叶可直接食用的天然绿色有机植物,并且全株可入药[3].明日叶拥有独特的芳香气味,用水焯过其茎叶后适口性增加,可用于熬汤、煸食、煨食或凉拌.明日叶茎叶含有人体所需的多种维生素及其他植物没有的B12、氨基酸、矿物质和微量元素,还含有抗氧化的查尔酮[4]、类黄酮[5]、香豆素[6]、有机锗等天然活性成分,具有抗肿瘤、抗艾滋病、抗衰老、抗溃疡、抗血栓、抗过敏、降血脂、降低胆固醇等功效[7],是促进身体健康、强健体魄、祛病延年的全营养食品,在日本及国内部分市场少量高价供应,具有较高的经济应用价值及广阔的开发前景.
随着人们近年来对明日叶保健功能的认识逐渐深入,明日叶作为一种强身健体、延年益寿的蔬菜被引入我国[3],目前在台湾、上海、海南、山东、贵州等地均有种植.明日叶良种的需求量伴随着种植面积逐渐扩大而增长.然而,生产上播种用的明日叶种子发芽十分迟缓且参差不齐,发芽率一般仅有5%~20%,严重影响其栽培生产.发芽实验可以检测种子的萌发潜力,评估其田间种植价值.Zhao et al[8]发现种子休眠类型对种子初萌发时间有显著影响.Li et al[9]发现低浓度(10 mol·L-1)的赤霉素(GA3)提高了毛竹种子的发芽率、发芽势、活力指数和呼吸速率,从而促进了毛竹种子的萌发,而高浓度(50 mol·L-1)的GA3抑制了种子的萌发.低浓度GA3加速了淀粉和脂肪的分解,促进了细胞液泡的形成,而高浓度GA3破坏了细胞器,增加了细胞的内吞作用.Lv et al[10]发现破坏褪黑素生物合成酶基因血清素n⁃乙酰基转移酶(SNAT)或n⁃乙酰羟色胺甲基转移酶(ASMT)可促进拟南芥种子萌发.因此,恰当正确的催芽办法对培养明日叶壮苗、提高其产量和品质十分重要.米永伟等[11]去除当归果翅后有效提高了种子发芽质量.索文龙等[12]发现低温及喷施乙烯对烟草种子萌发质量有所帮助.周红海和周欣悦[13]研究发现,水引发“两段浸种法”可增强早稻种子对抑制其生长作用的杀菌剂的抗性.崔少杰等[14]发现0.30%的KNO3溶液能够有效促进草莓种子萌发.KNO3溶液不仅能诱导植物体内生长激素的合成,调控其活力,还能消除萌发抑制物对种子的影响,使种子活力得到提高,促进发芽.本研究借鉴上述植物催芽处理方法,特别是近缘蔬菜的处理方法和研究结果,采用温水浸种、KNO3溶液浸种、不同植物激素浸种和不同强氧化剂浸种的方法处理明日叶种子,研究其种子发芽特性以及有效的催芽方法,为提高明日叶种子的发芽率,提高明日叶的产量和经济收益奠定基础.
1 材料与方法
1.1 供试种子
本实验测试的种子系2021年11月从山东青岛购买的当年生产的绿茎明日叶种子,其种子形态饱满,色泽新鲜.明日叶种子为双悬果,整体呈纺锤形,顶端略尖,底部呈扁圆形,种皮表面有棱,颜色为黄绿色.称量计算得该批明日叶种子的千粒重为15.78 g,平均长度为1.02 cm,平均宽度为0.51 cm.购置后的种子置于冰箱在4 ℃条件下暂时保存.
1.2 试剂与设备
本实验使用的试剂:KNO3,南京晚晴化玻仪器有限公司;GA3(赤霉素,上海迈瑞尔化学技术有限公司);NAA (α⁃萘乙酸,上海柏卡化学技术有限公司);SA(水杨酸,南京寿德生物科技有限公司);IAA(吲哚乙酸,南京寿德生物科技有限公司);6⁃BA (6⁃苄氨基嘌呤,江苏艾康生物医药研发有限公司);KMnO4,南京晚晴化玻仪器有限公司;30% H2O2溶液,南京晚晴化玻仪器有限公司;8% NaClO溶液,南京寿德生物科技有限公司.人工气候箱(QHX⁃40085⁃Ⅲ)为上海新苗医疗器械制造有限公司生产,另有滤纸、镊子、培养皿等.
1.3 浸种催芽处理
本实验自2021年12月开始,在南京大学生命科学学院植物信息与生态实验室人工气候箱(QHX⁃40085⁃Ⅲ)内进行.清水洗净所有种子,捞出后自然风干其表面水分至松散分开,然后进行浸种催芽实验.共设计四个种子发芽实验,以25 ℃蒸馏水浸种12 h后放入25 ℃黑暗条件下催芽为对照组,每个发芽实验设置不同处理,每个处理三次重复,完全随机排列,每次重复100粒种子(表1).参考以往相关研究方法,特别是伞形科植物种子萌发的浸种催芽等处理方法,设置以下条件对种子萌发进行探究:清水[15]、KNO3溶液[16]、植物激素[17-20]处理浸种时间为12 h[21],强氧化剂[22-23]处理浸种时间为20 min[24],浸种后用清水洗净,均匀放置于有滤纸的培养皿中,将滤纸润湿后置于适宜条件下进行催芽实验(表2,表3).
表1 明日叶种子的不同温水浸种处理
Table 1
| 浸种温度(℃) | 催芽温度(℃) | 催芽光照条件(光照强度单位:lx) |
|---|---|---|
| 25 (Control) | 25 (Control) | 0 |
| 45 | 15,22,30 | 0 |
| 55 | 15,22,30 | 0 |
| 65 | 15,22,30 | 0 |
| 55 | 15 | 0 |
| 15 | 3000 | |
| 15 | 催芽处理后直接播种进土壤 |
表2 明日叶种子的KNO3溶液及五种植物激素浸种处理
Table 2
| 处理 | 浓度(mg·L-1) |
|---|---|
| KNO3溶液 | 0,10,50,100,250,500,750,1000 |
| GA3溶液 | 0,50,100,200,300,400 |
| NAA溶液 | 0,50,100,200,300 |
| SA溶液 | 0,100,200,300,400 |
| IAA溶液 | 0,20,50,100,200 |
| 6⁃BA溶液 | 0,5,10,20,30,50,100,200,300 |
表3 明日叶种子的三种强氧化剂催芽处理
Table 3
| 处理 | 浓度(mg·L-1) | 浸种时间 |
|---|---|---|
| Control | 0 | 12 h |
| KMnO4溶液 | 100,250,500,1000,3000 | 20 min |
| H2O2溶液 | 3×103,1×104,3×104,5×104, 1×105,3×105 | 20 min |
| NaClO溶液 | 100,1000,10000 | 20 min |
1.4 指标测量及计算
每24 h补充水分以保持滤纸湿润并统计一次当天发芽数.当胚芽达到种子一半长时即为发芽,以连续3 d不再发芽为发芽率统计时间.发芽实验结束时,用软尺测定明日叶幼苗的根长、下胚轴长,计算如下[25]:
其中,Ft 表示在第t日时的累计发芽数,Vt 表示第t日的种子发芽数,Rt 表示第t日时的累计烂种数,DT 表示第T日,NT 表示供试种子总数,F23表示种子萌发第23 d时累计种子发芽数.
1.5 数据分析
采用Microsoft Excel 2016进行数据记录整理,IBM SPSS Statistics 23.0进行单因素ANOVA方差分析及Duncan多重比较(α=0.05),用Graphpad prism 8.0绘图.
2 结果与分析
2.1 不同浸种温度和发芽温度对明日叶种子发芽的影响
如图1所示,在三个不同的浸种温度条件下(浸种时间均为12 h),当浸种温度为55 ℃时,明日叶种子的发芽率、发芽势和发芽指数均高于浸种温度为45 ℃和65 ℃这两个处理组.在相同浸种温度条件下,随着发芽温度的升高,明日叶种子35 d后的发芽率、发芽势和发芽指数显著下降(p<0.05).而烂种率随着发芽温度的提高而提高,当发芽温度为30 ℃时,烂种率显著最高,分别为53.85%,51.9%和62.5%.以上结果表明,55 ℃的温水浸种是使种子的发芽率、发芽势和发芽指数最优的浸种温度.
图1
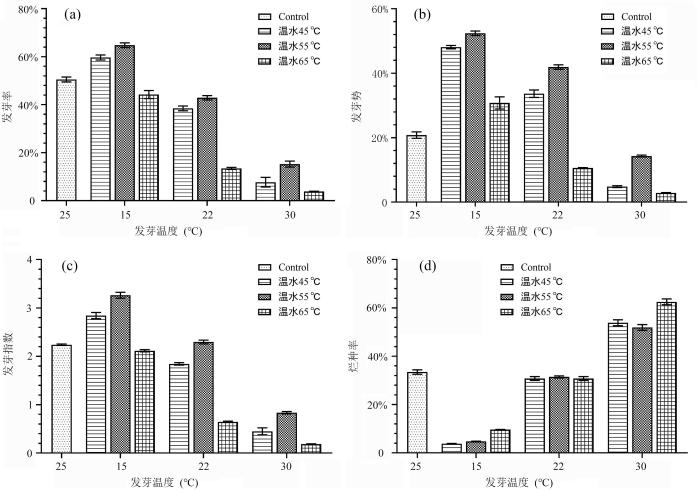
图1
不同浸种温度及发芽温度对明日叶种子发芽的影响
Fig.1
Influence of different soaking temperatures and germination temperatures on germination of Angelica keiskei seeds
2.2 温水浸种处理对明日叶种子萌发的影响
明日叶种子经冷水(对照组,25 ℃)浸种12 h,第19 d开始萌发,温水(55 ℃)浸种12 h并在黑暗条件下催芽,第16 d开始萌发.但从35 d后的发芽结果看,温水浸种(光照组)发芽率最高,达到63%,冷水浸种(对照组)的只有50.5%,浸种后直播的发芽率仅有20%,显著低于冷水浸种(对照组)及其他两种处理(p<0.05).上述结果表明,明日叶种子经温水浸种(55 ℃)处理12 h后,在光照条件下发芽的处理组表现最优.
2.3 不同浓度KNO3溶液处理对明日叶种子萌发的影响
本实验通过设置不同浓度梯度的KNO3处理,分析明日叶种子的发芽进程、发芽率、发芽势、发芽指数等指标,探究不同KNO3浓度对明日叶种子发芽的影响,从而获得最佳浓度KNO3溶液处理的催芽方式.
通过观察不同浓度KNO3处理的种子发芽情况,从第15 d逐日统计种子发芽数,第35 d发芽结束,发现不同浓度KNO3处理组,第18 d有少量种子发芽,第21 d左右萌发数量明显增多,第30 d后种子发芽趋势逐渐变缓.
如图2a所示,不同浓度KNO3处理种子的发芽率无显著差异,但均显著高于对照组(p<0.05),提高了43.74%~65.98%.
图2

图2
不同浓度KNO3溶液处理对明日叶种子发芽的影响
Fig.2
Effect of KNO3 solution at different concentrations on germination of Angelica keiskei seeds
如图2b所示,在所有处理组中,随着KNO3溶液浓度的提高,明日叶种子的发芽势呈先上升后下降的趋势,当KNO3浓度达到750 mg·L-1,发芽势最高,为59.56%,比对照组提高了186.48%.
如图2c所示,不同浓度KNO3溶液处理期间明日叶种子的发芽指数无显著差异(p>0.05),比对照组提高了101.84%~150.85%.在KNO3溶液浓度上升后并未对明日叶种子的发芽率及发芽指数造成显著影响,而发芽势在KNO3溶液浓度为750 mg·L-1时表现最优.
下胚轴指从子叶着生处以下生出的最初茎的部分.如图3所示,一定浓度的KNO3溶液处理促进了明日叶幼苗的下胚轴长,当KNO3溶液浓度大于500 mg·L-1时,下胚轴长被显著抑制(p<0.05).明日叶幼苗根长随着KNO3溶液浓度的升高呈现先上升后下降趋势.其中,100 mg·L-1 KNO3溶液处理的明日叶幼苗根长最长,达到2.48 cm,比对照组提高了64.24%.
图3

图3
不同浓度KNO3溶液处理对明日叶幼苗生长的影响
Fig.3
Effects of KNO3 solution treatment at different concentrations on the growth of Angelica keiskei seedlings
2.4 不同植物激素处理对明日叶种子萌发的影响
植物激素可以调控植物的生长发育进程,是其体内产生的一些含量极低却可以调节自身生理过程的有机化合物[26].本研究使用不同浓度梯度的植物激素如GA3,NAA,IAA,SA,6⁃BA对明日叶种子进行浸种处理,分析其发芽进程、发芽率、发芽势、发芽指数等指标,探究不同植物激素对明日叶种子发芽的影响,从而获得最佳浓度植物激素处理的催芽方式.
2.4.1 不同植物激素处理对明日叶种子发芽进程的影响
观察不同浓度GA3处理的种子发芽情况,从第16 d起(开始发芽)逐日统计种子发芽数,第35 d发芽结束.发现不同浓度GA3处理后的种子发芽率均显著高于对照组(p<0.05).在第17 d,不同浓度GA3处理组均有少量种子发芽,第22 d左右萌发数量明显增加,第30 d后种子发芽趋势逐渐变缓.不同浓度NAA处理的种子前期发芽缓慢甚至不发芽,和对照组一样约在第18-19 d时发芽.第23 d左右发芽速度达到顶峰,随后逐渐变缓.浓度为50,100和200 mg·L-1 NAA处理的种子的发芽率均高于对照组,300 mg·L-1 NAA处理的种子发芽趋势和发芽率显著低于对照组和其他处理组(p<0.05).不同浓度SA处理的明日叶种子早于其他激素处理组开始萌发(第13 d),并且都于约第31 d停止萌发.虽然发芽速度显著高于对照组和其他植物激素处理组(p<0.05),但最后的发芽率均为~60%,低于NAA和GA3处理组.IAA处理组的发芽高峰较早,约在第19 d发芽达到顶峰,后续发芽数量趋于平稳.其中只有100 mg·L-1处理的发芽率显著低于对照组(p<0.05),并于第28 d就发芽完毕,后续没有再发芽.6⁃BA (6⁃苄氨基嘌呤)是一种细胞分裂素,100 mg·L-1 6⁃BA 处理组在第23 d才开始萌发,第28 d停止萌发,最终发芽率仅为6.52%.
2.4.2 不同植物激素处理对明日叶种子发芽率的影响
如图4a所示,在设置的浓度梯度范围内,GA3处理过的种子发芽率呈先上升后下降的趋势,其中100 mg·L-1 GA3处理的种子发芽率最高,为81.19%,发芽率最低的是400 mg·L-1 GA3处理组,仅41.58%,比对照组低了17.66%.在NAA处理组中,100和200 mg·L-1 NAA处理组的发芽率分别比对照组高了37.84%和26.20%;50和300 mg·L-1 NAA处理组与对照组相比无显著差异(p>0.05).低浓度6⁃BA处理的种子发芽率无显著差异(p>0.05),但都显著高于对照组(p<0.05),6⁃BA浓度高于100 mg·L-1后发芽率显著降低,200和300 mg·L-1 6⁃BA处理组的种子无萌发现象.在设置的浓度梯度范围内,IAA处理组种子发芽率随着浓度升高呈先下降后上升趋势,其中20 mg·L-1 IAA处理的种子发芽率显著高于其他处理组(p<0.05),为71.57%.SA浓度在100~400 mg·L-1时,种子的发芽率无显著差异(p>0.05).
图4

图4
五种植物激素及其不同浓度梯度处理对明日叶种子发芽的影响
Fig.4
Effects of five plant hormones and their different concentration gradients on germination of Angelica keiskei seeds
2.4.3 不同植物激素处理对明日叶种子发芽势的影响
发芽势可以表征种子质量好坏,是指在发芽时日发芽种子数达到最多时,萌发的种子数占样品种子总数的百分率.在发芽率一定时,发芽势越高,种子生命力越强.若发芽率较高但发芽势低则代表出苗不齐、弱苗多[24].如图4b所示,在GA3处理组中,100 mg·L-1处理组的发芽势最高,为31.68%,比对照组提高了52.38%,50 mg·L-1 GA3处理组的发芽势最低,仅为9.8%,其余处理组与对照组相比无显著差异(p>0.05).NAA处理组中,100和200 mg·L-1 NAA处理组的发芽势相同,比对照组高了22.61%,50 mg·L-1 NAA处理组与对照组相比无显著差异(p>0.05).6⁃BA浓度在5~10 mg·L-1时,发芽势随着浓度的提高呈现递增趋势,当浓度大于10 mg·L-1时,随着6⁃BA浓度的增加,发芽势呈下降趋势.第23 d时,200和300 mg·L-1 6⁃BA处理组仍无萌发迹象,故无法测量得到发芽势(图4b).在设置的浓度梯度范围内,IAA处理过的种子发芽势呈先下降后上升趋势,其中20 mg·L-1 IAA处理的明日叶种子发芽势显著高于对照组与其他IAA处理组(p<0.05),为44.12%.虽然SA浓度在100~400 mg·L-1时,明日叶种子的发芽率无显著差异(p>0.05),但发芽势随着浓度的增加呈下降趋势,其中100 mg·L-1SA处理过的明日叶种子发芽势最高,为45.53%.
2.4.4 不同植物激素处理对明日叶种子发芽指数的影响
发芽指数是反映种子活力高低的指标之一.发芽指数越高,表征种子活力越高[24].经过植物激素处理过的种子的发芽指数集中在1.8~3.4.GA3浓度为100 mg·L-1时明日叶的发芽指数最高,为3.3724,比对照组提高了51.26%.NAA浓度为100 mg·L-1时,发芽指数最高,为2.8033,比对照组提高了25.73%.6⁃BA浓度为20 mg·L-1时,发芽指数最高,为3.1596,比对照组提高了41.71%.如图4c所示,在设置的浓度梯度范围内,GA3,NAA,6⁃BA处理组的发芽指数随着浓度的增加呈先上升后下降趋势.IAA浓度为20 mg·L-1时发芽指数最高,为3.4366,比对照组提高了54.14%.SA浓度为100 mg·L-1时,发芽指数最高,为3.0140,比对照组提高了35.18%.在设置的浓度梯度范围内,IAA,SA处理组的发芽指数随着浓度的增加呈先下降后上升的趋势.
2.4.5 不同植物激素处理对明日叶幼苗生长的影响
如图5所示,在GA3,6⁃BA处理组中,明日叶幼苗的下胚轴长和根长均随着浓度的增加呈先上升后下降趋势.GA3浓度为100 mg·L-1时下胚轴最长,达到2.41 cm;GA3浓度为200 mg·L-1时,根系最长,达到2.30 cm.在NAA,IAA,SA处理组中,明日叶幼苗的下胚轴长和根长均随着浓度的增加呈下降趋势.除50 mg·L-1 NAA溶液处理以外,其余浓度NAA处理的下胚轴长与对照组相比无显著差异(p>0.05),但100 mg·L-1NAA溶液处理显著促进根系的生长(p<0.05).而200 mg·L-1 IAA,300和400 mg·L-1 SA处理均显著抑制幼苗的下胚轴伸长(p<0.05).
图5

图5
不同植物激素处理对明日叶幼苗生长的影响
Fig.5
Effects of different plant hormone treatments on the growth of Angelica keiskei seedlings
2.5 不同强氧化剂处理对明日叶种子萌发的影响
强氧化剂可以通过对种子表皮灭菌,提高种皮透性,促进种内抑制物质渗出等方式促进种子萌发[27].本实验通过设置不同浓度梯度的强氧化剂,包括NaClO,KMnO4和H2O2,分析明日叶种子的发芽进程、发芽率、发芽势、发芽指数等指标,探究不同浓度强氧化剂对种子发芽的影响,从而获得最佳浓度强氧化剂处理的催芽方式.
2.5.1 不同强氧化剂处理对明日叶种子发芽进程的影响
通过观察不同的浓度NaClO处理的种子发芽情况,发现发芽启动时间大大提前,约在第9 d时开始萌发.不同浓度的NaClO浸泡不同时间后的明日叶种子在第20 d左右萌发数量明显增加,在27 d左右达到顶峰,随后趋于平缓.KMnO4处理组从第16 d开始萌发,第20 d后萌发数量明显增多,第35 d停止萌发.H2O2处理组中,3×105 mg·L-1浓度处理的种子第25 d才开始萌发,发芽速度缓慢,最终发芽率显著低于对照组,3×104,5×104和1×105 mg·L-1浓度处理过的种子前期萌发速率缓慢,低于对照组,在第28 d之后萌发数量明显增加.
2.5.2 不同强氧化剂处理对明日叶种子发芽率的影响
如图6a所示,在一定范围内随着KMnO4浓度的提高,明日叶种子的发芽率呈上升趋势,其中500 mg·L-1处理的发芽率最高,为84.92%,比对照组提高了68.16%.H2O2处理组中,3×105 mg·L-1浓度处理的发芽率仅为25%,比对照组低了50.5%;1×105 mg·L-1 H2O2处理的种子发芽率最高,为77.66%,比对照组提高了53.78%.在NaClO处理组中,不同浓度处理组间的发芽率无显著差异(p>0.05).
图6

图6
不同强氧化剂及其不同浓度处理对明日叶种子发芽的影响
Fig.6
Effects of different strong oxidants and their different concentrations on germination of Angelica keiskei seeds
2.5.3 不同强氧化剂处理对明日叶种子发芽势的影响
如图6b所示,KMnO4处理组中,100和500 mg·L-1 KMnO4处理组的发芽势分别比对照组高了92.4%和121.4%,其余处理组间无显著差异(p>0.05).在一定浓度范围内,H2O2处理过的种子的发芽势随浓度的增大呈先下降后上升的趋势,其中3×103 mg·L-1 H2O2处理组的发芽势最高,为33.01%,比对照组提高了58.78%;3×105 mg·L-1 H2O2处理组的种子在统计当天(第23 d)并无发芽,故无法统计到发芽势.在NaClO处理组中,100和1000 mg·L-1两组无显著差异(p>0.05),分别比对照组高了91.01%和85.52%.
2.5.4 不同强氧化剂处理对明日叶种子发芽指数的影响
如图6c所示,KMnO4处理组中,浓度范围为250~3000 mg·L-1时,发芽指数无显著差异(p>0.05),但均高于对照组.3×103~1×105 mg·L-1的H2O2浸种处理后,对种子发芽指数的影响未达到显著水平(p>0.05),3×105 mg·L-1 H2O2处理组的发芽指数仅为0.7916,比对照组低了64.50%.在NaClO处理组中,10000 mg·L-1处理组的发芽指数显著高于对照组(p<0.05),提高了66.97%.
2.5.5 不同强氧化剂处理对明日叶幼苗生长的影响
如图7所示,100和250 mg·L-1 KMnO4处理组的下胚轴显著伸长(p<0.05),但根长无明显变化(p>0.05).其余KMnO4处理组与对照组相比下胚轴长无显著差异,但1000和3000 mg·L-1 KMnO4处理组的根长显著减小(p<0.05).在H2O2处理组中,3×104 mg·L-1H2O2处理的下胚轴最长,达到1.90 cm,比对照组提高了58.33%.随着H2O2溶液浓度的升高,下胚轴长被显著抑制(p<0.05).3×105 mg·L-1 H2O2处理组的根长显著减小,比对照组短了58.33% (p<0.05).在NaClO处理组中,10000 mg·L-1处理组显著抑制了下胚轴及根的长度(p<0.05).
图7

图7
不同强氧化剂处理对明日叶幼苗生长的影响
Fig.7
Effects of different strong oxidant treatments on the growth of Angelica keiskei seedlings
3 讨论
3.1 适宜的浸种温度明显缩短明日叶种子的发芽进程
温度可能通过影响种子内部与萌发相关的酶的活性,从而调控种子的萌发进程[30].我们的研究发现较低的发芽温度(15 ℃)更有利于明日叶种子的萌发,梁润芳等[31]对羊草种子的研究表明,低温处理可明显提高乌珠穆沁羊草种子和长岭羊草种子的发芽率、发芽势、发芽速率、发芽指数、活力指数和苗长.可能是由于较低的温度可以激活种子内部的化学物质,并在一定水平上抑制霉菌的滋生.而高温(30 ℃)发芽处理的种子,由于种子呼吸加强或导致种胚内部产生变化而抑制萌发,其发芽率、发芽势、发芽指数显著降低,大部分种子出现烂种现象.王楠等[15]研究了同属植物当归(Angelica sinensis)在15和25 ℃条件下种子的发芽率,发现25 ℃发芽率高于15 ℃.这与本文得出最优发芽温度为15 ℃的研究结果有一定差异,可能是由于浸种温度的不同导致的.本实验还探究了浸种后处理对种子萌发特性的影响,发现在浸种后置于培养皿中发芽的种子无论在光照还是黑暗条件下发芽率均有所提高;相反,温水浸种后直播的种子发芽率仅有20%,显著低于对照组及其他两个处理组(p<0.05),推测是由于种子埋在土中,更容易受土中微生物的干扰而失去活力.
3.2 适宜浓度的KNO3显著提高明日叶种子的发芽势
不同浓度KNO3处理间种子的发芽率和发芽指数无显著差异,但均显著高于对照组(p<0.05).但对发芽势来说,在一定范围内,随着KNO3浓度的提高,明日叶种子的发芽势呈先上升后下降的趋势,当KNO3浓度达到750 mg·L-1,发芽势最高,为59.56%,比对照组提高了186.48%.这与Thongtip et al[25]发现的0.4% KNO3溶液可提高圣罗勒种子的发芽率和萌发指数、降低平均萌发时间的结果相似.Rizk et al[32]发现250和500 mg·L-1 KNO3溶液能显著刺激反枝苋种子生长,提高其胚根长度,而本实验中100 mg·L-1 KNO3溶液处理显著促进了明日叶幼苗的下胚轴长及根长,但当KNO3溶液浓度大于500 mg·L-1时,幼苗生长被显著抑制.实验结果表明,硝酸钾等盐溶液可能会通过影响种子内部的化学物质代谢来影响种子发芽进程及幼苗生长.刘镎等[33]关于硼酸和硝酸钾引发甜菜种子的研究中有类似的报道.
3.3 适宜的外源植物激素添加可促进明日叶种子萌发
植物激素对明日叶种子的发芽率、发芽势、发芽指数均有促进作用,其中100 mg·L-1 GA3溶液处理后发芽率最高,达到81.19%.这与Wang et al[34]发现100 mg·L-1 GA3溶液处理后,栽培樱桃、野生樱桃和甜樱桃种子的发芽率分别达到最大值64%,24%和40%的研究结果一致.但本研究中GA3溶液的最佳浓度范围与前人在其他种子上的研究结果存在一定差异:Hosseini et al[35]证明0.1% GA3溶液处理对番石榴种子萌发和幼苗生长均有促进作用;王宁等[17]发现节麦种子经500 mg·L-1 GA3溶液处理后发芽率和发芽势得到显著提高;殷武平等[36]的研究结果表明用250 mg·L-1的GA3溶液浸种芹菜种子的效果最佳.推测是由于不同植物种子种皮通透性及内源物质不同,导致GA3对种子萌发的促进作用也不同.本实验中,100 mg·L-1 GA3溶液处理显著促进明日叶茎伸长,200 mg·L-1 GA3显著促进明日叶根系生长,这与殷武平等[36]的研究结果类似.本实验进一步证明,适宜浓度的GA3溶液能够在一定程度上加速细胞分裂,促进细胞分化,有利于促进伞形科植株幼苗生长,在农业生产上可以提高植物的产量.
3.4 适宜的强氧化剂浓度可促进明日叶种子萌发
4 结论
本研究针对实际生产中不同的处理方式,组内对比筛选出四种催芽效果最佳处理,分别为温水55 ℃浸种后在光照条件下15 ℃发芽处理;100 mg·L-1 GA3溶液处理;750 mg·L-1 KNO3溶液处理;500 mg·L-1 KMnO4溶液处理.通过综合比较这四种情况下明日叶种子的发芽率、发芽势、发芽指数、下胚轴长及根长,考虑到外源添加药剂的可获得性,推荐使用100 mg·L-1 GA3溶液(25 ℃浸种12 h后放入黑暗环境,25 ℃条件下催芽)进行明日叶种子催芽,可达到最佳效果.
参考文献
Prenylated chalcones 4⁃hydroxyderricin and xanthoangelol stimulate glucose uptake in skeletal muscle cells by inducing GLUT4 translocation
明日叶的组织培养与快速繁殖
Tissue culture and rapid propagation of Angelica keiskei Koidz
Non⁃deep simple morphophysiological dormancy in seeds of Angelica keiskei (Apiaceae)
Antidiabetic activities of chalcones isolated from a Japanese herb,Angelica keiskei
Chalcones from Angelica keiskei (ashitaba) inhibit key Zika virus replication proteins
Recent advances on anticancer activity of coumarin derivatives
Angiotensin⁃converting enzyme inhibitors from plants:A review of their diversity,modes of action,prospects,and concerns in the management of diabetes⁃centric complications
Germination characteristics is more associated with phylogeny⁃related traits of species in a salinized grassland of northeastern China
Ultrastructure change and transcriptome analysis of GA3 treatment on seed germination of moso bamboo (Phyllostachys edulis)
Melatonin inhibits seed germination by crosstalk with abscisic acid,gibberellin,and auxin in Arabidopsis
当归果翅对种子吸水与发芽进程的影响
Responses of water absorption and germination of Angelica sinensis seeds to winged perianth removal
外源植物生长调节剂对不同发育天数烟草种子萌发的影响
Effects of exogenous plant growth regulators on seed germination of tobacco at different develop⁃mental days
水引发条件下“两段浸种法”对早稻种子发芽率和秧苗素质的影响
Study of hydropriming on rice seed germination rate and seedling quality by using two⁃phase soaking method
不同药剂处理对草莓种子萌发的影响
Effects of different regents treatments on the germination of strawberry seeds
当归种子活力
Seed germination vigor of Angelica sinensis
打破野生藏沙蒿种子休眠方法的筛选与评价
Screening and evaluation of methods for breaking seed dormancy of wild Artemisia wellbyi
GA3浸种对入侵植物节节麦种子破眠及发芽特性的影响
Effectiveness of exogenous GA3 for dormancy breaking in invasive Aegilops tauschii,and effect on germination physiology
外源生长物质对干旱胁迫条件下甜高粱种子萌发的影响
水杨酸引发对红麻幼苗耐盐性的影响
Effect of salicylic acid priming on salt tolerance of Kenaf seedlings
不同浓度6⁃BA包衣芹菜种子对萌发效果的影响
Effect of different concentration 6⁃BA coated celery seed on germination
种子引发对芹菜种子发芽、幼苗生长及产量的影响
The effect of seed priming on celery seed germination,seedling growth and yield
次氯酸钠处理种子对芹菜幼苗可溶性糖含量及相关基因表达的影响
Effects of seed⁃soaking with sodium hypochlorite on soluble sugar content and expression profiles of related genes of celery seedlings
Promotion of seed germination and early plant growth by KNO3 and light spectra in Ocimum tenuiflorum using a plant factory
Transport mechanisms of plant hormones
不同消毒剂对濒危药材西藏龙胆种子的消毒效果及萌发的影响
Effects of different disinfectants on sterilization and germination of Gentiana tibetica King ex Hook
.
A perspective on secondary seed dormancy in Arabidopsis thaliana
不同处理方法对紫花苜蓿种子萌发的影响
Effect of different treatment methods on alfalfa seed germination
Temperature during seed maturation controls seed vigour through ABA breakdown in the endosperm and causes a passive effect on DOG1 mRNA levels during entry into quiescence
低温和光照对羊草种子萌发的影响
Effects of low⁃temperature and light on seed germination of Leymus chinensis
Breaking dormancy and enhancing germination of Avena sterilis L. and Amaranthus retroflexus L. weeds by gibberellic acid and potassium nitrate to keep soil and crops healthy
硼酸和硝酸钾引发甜菜种子的研究
Study on seed initiation of sugar beet by boric acid and potassium nitrate
Effect of gibberellin treatment on dormancy⁃breaking and germination of cherry Seeds
Improving seed germination and seedling growth of guava under heat and osmotic stresses by chemical and hormonal seed treatments
不同浸种和催芽处理对芹菜种子发芽的影响
Effects of different seed soaking and germination treatments on the germination of celery seeds
Mechanism of sugarbeet seed germination enhanced by hydrogen peroxide

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}